Die Biomechanik der Morphogenese
Forschungsbericht (importiert) 2013 - MPI für Pflanzenzüchtungsforschung
Die Biomechanik der Morphogenese
In den letzten Jahren hat sich unser Verständnis von der Pflanzenmorphogenese auf molekularer Ebene enorm erweitert. Wir verfügen heute über Erkenntnisse darüber, wie Genexpression und interzelluläre Signaltransduktion molekulare Netzwerke steuern, die sich auf die Zelldifferenzierung auswirken. Viele der an der Steuerung der Pflanzenorganform beteiligten Gene sind inzwischen bekannt. Anders als bei Tieren können Pflanzenzellen sich nicht im Organismus bewegen, sondern sind starr aneinander gebunden. Dadurch sind die für die Pflanze zur Formausbildung zur Verfügung stehenden Mechanismen begrenzt. Ohne bewegliche Zellen muss sich die Morphogenese zur Entwicklung verschiedener Formen auf differentielles Wachstum stützen. Und obwohl die Form nur auf Gewebeniveau entsteht [1], ist sie das Ergebnis von sorgfältig koordinierten Reaktionen einzelner Zellen. Deshalb ist das Verstehen des Wachstumsverhaltens auf zellulärer Ebene grundlegend für das Verstehen der Morphogenese.
Pflanzenzellen sind von einer starren Zellwand umschlossen, die den Zellinnendruck (Turgor) hält. Dieser Druck ist groß und kann sogar noch weit über dem Druck eines Autoreifens liegen. Trotz dieser enorm hohen Kraft können sich Zellen auf mehr als das Tausendfache ausdehnen, ohne zu platzen. Wie wird diese mechanische Meisterleistung erreicht? Es wird angenommen, dass die Mechanik der Zellwandausdehnung über eine regulierte Zellwandlockerung abläuft, was jedoch bislang nur in Ansätzen untersucht ist. Allerdings konnten viele Strukturbestandteile der Zellwand und auch mehrere Enzyme identifiziert werden, die diese modifizieren [2]. Letztlich müssen die an der Formregulierung beteiligten genetischen Netzwerke und Signalnetze genau in diese Bestandteile eingreifen, um die physikalischen Eigenschaften der Zellwand derart zu verändern, damit sie wachsen kann.
Die Messung der physikalischen Eigenschaften von Pflanzenzellen
Die Mechanik der wachsenden Zellen wird mit Hilfe einer mikro-robotischen Technik untersucht, die wir als Zellenkraftmikroskopie - Cellular Force Microscopy (CFM) -bezeichnen. Diese Technik haben wir in Zusammenarbeit mit dem Nelson-Labor der ETH Zürich entwickelt [3]. Mit Hilfe der CFM können die physikalischen Kräfte in Pflanzenzellen durch Indentation der Zelle mit einer dünnen Sonde, die mit einem Kraftsensor verbunden ist, direkt gemessen werden. Die ermittelten Kraft- und Verschiebungsdaten werden anschließend zur Berechnung der Steifigkeit der jeweiligen Probe verwendet. Ähnlich wie die Rasterkraftmikroskopie (Atomic Force Microscopy – AFM) arbeitet auch die CFM mit computergesteuerten Aktoren, die die Sonde bewegen. Mit Hilfe dieser Technik entstehen hochauflösende Steifigkeits- und Höhendarstellungen. Abbildung 1 zeigt eine Steifigkeitsdarstellung der CFM-Indentation in Epidermiszellen der Zwiebel. Bei prallen Zellen erscheint das Zentrum der Zelle steifer als die Querwände.
. Die blauen Quadrate zeigen weiche Areale, die roten steifere Areale an. Farbbalken in N/m2, Maßstabsbalken 20 µm.")
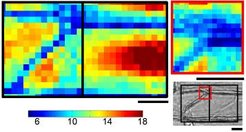
Die CFM ist besonders für die Messung von Kräften in Pflanzen geeignet. Diese Technik schließt nämlich die Lücke zwischen den sehr geringen Kräften, die mit AFM gemessen werden, und den wesentlich stärkeren Kräften, die durch traditionelle Wägezellen gemessen werden. CFM bietet zudem eine ausgezeichnete Geometriefreiheit. Sie ermöglicht große Bewegungen bis hin zu Zentimetern und verfügt über optisch getrackte Closed-Loop-Positionierer, die nanometergenaue Bewegungen durchführen können. Mit der langen, schlanken Sonde werden außerdem Bereiche zugänglich, die mit dem AFM-Arm unerreichbar wären, und es lassen sich Einzelzellenablationen mittels mikrochirurgischer Experimente durchführen. Eine eigens von den Forschern entwickelte, anwendungsspezifische Steuerungssoftware ermöglicht es zudem, das System für Einzelexperimente individuell anzupassen. So wurde beispielsweise kürzlich eine Miniaturversion des Systems entwickelt, die in einem senkrechten Konfokal-Mikroskop verwendet werden kann (Abb. 2). Das neuartige Konfokal-CFM ermöglicht eine quasi-simultane Messung von Genexpression und physikalischen Kräften auf zellulärer Ebene.


Wirken mechanische Kräfte auf die Genexpression zurück?
Es gibt immer mehr Beweise dafür, dass mechanischer Stress auf Hormonsignalwege und das Wachstum zurückwirken und bei der Musterbildung im Entwicklungsprozess eine zentrale Rolle spielen könnten. In welchem Umfang sich die physikalischen Eigenschaften von Zellen und ihr lokales Umfeld auf Genexpression und Morphogenese auswirken, ist derzeit Gegenstand intensiver Forschung. Eines der offensichtlichen Beispiele für dieses Phänomen ist die Verdickung der Knochen als Reaktion auf Beanspruchung. Wenn wir unsere Knochen durch Heben von Gewichten oder durch anstrengende Aktivitäten belasten, nimmt ihre Dichte zu. In Pflanzen erfolgt eine ähnliche Reaktion. Stängel verstärken sich als Reaktion auf Belastungen durch Wind – ein Phänomen, das sich im Labor durch Anwendung künstlicher Belastung reproduzieren lässt. Tiefergreifende Beispiele gibt es in Tiersystemen. So hat man etwa herausgefunden, dass Differenzierungsentscheidungen von Stammzellen von der Steifigkeit des Substrats abhängen, auf dem sie kultiviert werden [4]. Mesenchymale Stammzellen, die auf sehr weichem Substrat platziert werden, exprimieren genetische Marker mit Gehirnzellenidentität, während ein steiferes Medium Muskelzellen-Marker induziert. Und auf einem sehr starren Substrat werden Marker mit Knochenzellenidentität beobachtet. In diesem System scheint die Substratsteifigkeit als Morphogen zu agieren, das die Zellen dazu veranlasst, ihre Schicksale in einem Gradient abhängigem Muster zu wählen. Könnten solche Mechanismen auch in Pflanzen existieren?
Eine Untersuchung der Stammzellnische in der Tomatentriebspitze scheint nahezulegen, dass Pflanzenzellen bei der Wahl oder Aufrechterhaltung ihrer Identität auch physikalische Signale nutzen. Pflanzenstammzellen befinden sich in Strukturen, die als Meristeme bezeichnet werden. Das Meristem an der Triebspitze ist für die Anlage aller flächigen Pflanzenteile zuständig. Die Stammzellen in der mittleren Zone der Triebspitze wachsen langsam und liefern Gründerzellen für den umliegenden Randbereich. Der Randbereich wiederum enthält schnell wachsende und sich teilende Zellen und ist derjenige Ort, an dem Blätter und Blüten entstehen. Alle Zellen des Meristems scheinen zwar physikalisch von gleicher Form, Größe und Zellwandstärke zu sein. Die verschiedenen Zonen weisen jedoch sehr unterschiedliche Wachstumsmuster auf und nur Zellen im Randbereich sind in der Lage, neue Organe zu bilden. Unterscheiden sie sich in ihren mechanischen Eigenschaften? Und wenn ja: Ist dieser Unterschied für die Entwicklung von Bedeutung? Zur Beantwortung dieser Fragen wurde der osmotische Druck experimentell verändert, um die Zellwandspannung in der Tomatentriebspitze zu manipulieren [5]. Mit Hilfe der von uns eigens entwickelten Software war es möglich, die Oberflächenverformung der Triebspitze als Reaktion auf Druckveränderungen nachzuvollziehen. Diese Ergebnisse wurden dann dazu verwendet, die Steifigkeit der Zellen in den peripheren und zentralen Zonen zu berechnen.
Zellen im schneller wachsenden Randbereich haben sich infolge zunehmender Zellwandspannung schneller ausgedehnt als Zellen in der langsamer wachsenden Stammzellnische. Das lässt vermuten, dass Zellen über die Fähigkeit verfügen müssen, sich elastisch zu verformen, um wachsen zu können, und dass die funktionelle Zonierung in der Triebspitze mechanisch definiert werden kann. Ob solche Unterschiede in den mechanischen Eigenschaften wiederum auf das Zellschicksal zurückwirken, wie im Tiersystem nachgewiesen, bleibt eine offene Frage, die es nun zu untersuchen gilt.
Verfolgen von Zellverformung und Wachstum in vivo
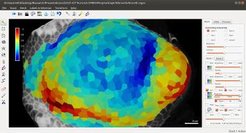
Damit Zellformveränderungen in vivo nachvollzogen werden können, wurde eine spezielle Bildgebungs- und Zellsegmentierungssoftware entwickelt: MorphoGraphX (Abb. 3). Da Pflanzen symplastisch wachsen - ihre Zellen sind über kleine Kanäle, sogenannte Plasmodesmata, miteinander verbunden - lassen sich durch Beobachtung der Formveränderung in der Oberflächenschicht von Zellen umfassende Informationen über die Morphogenese gewinnen. In vielen Systemen ist die Oberflächenschicht von Zellen jedoch nicht flach, weshalb Zellforminformationen dann verloren gehen, wenn konfokale Bilddaten auf eine Fläche projiziert werden. Dieses Problem wurde durch MorphoGraphX gelöst: Die Software ermöglicht eine Zeitraffer-Quantifizierung gekrümmter Oberflächenschichten von Zellen. Irgendwo zwischen 2 und 3D arbeitend, ist MorphoGraphX in der Lage, 3D-Konfokal-Bildstapel in Darstellungen gekrümmter Flächen umzuwandeln. Diese werden dann mit Hilfe von Standardalgorithmen weiter verarbeitet, die zweckentsprechend angepasst wurden. Neben der Verfolgung von zellgeometrischen Veränderungen kann die Software auch dazu eingesetzt werden, die Fluoreszenz genetischer Marker zu quantifizieren, sodass sich die Genexpression direkt mit der Zellformveränderung korrelieren lässt. Mit dieser Software wird inzwischen in vielen internationalen Labors gearbeitet.
. Der Farbbalken gibt das Vielfache der Zunahme des Zelloberflächenbereichs an.")
Wie wirkt sich die Zellproliferation auf Morphogenese und Wachstum aus?
Neben der Zellexpansion ist auch die Zellproliferation ein entscheidender Wachstumsfaktor. Die am schnellsten wachsenden Bereiche des Pflanzengewebes weisen oft hohe Zellteilungsraten auf. Eine Kernfrage ist, ob die Zellteilung schon allein das Wachstum anregt oder aber ob das Wachstum des Gewebes auf Organebene gesteuert wird und die Zellen einfach anfangen, sich zu teilen, sobald sie eine bestimmte Größe erreicht haben. Vor über 100 Jahren haben Errea und andere Wissenschaftler Standardregeln für die Zellteilung formuliert, die von Besson und Dumais [6] in einem Modell zusammengefasst wurden. Danach verhalten sich teilende Zellen wie Seifenblasen, wobei die Teilungswand eine Mindestenergiekonfiguration annimmt, die die Zelle in zwei gleiche Teile unterteilt. Mit einer solchen Hypothese der “kürzesten Wand” lassen sich viele Zellteilungsmuster für eine Vielzahl von Landpflanzen und Algen erklären. In Zusammenarbeit mit dem Labor von Weijers in Wageningen und dem Labor von Prusinkiewicz in Calgary haben wir eine 3D-Zeitreihe der frühen Embryonalentwicklung mit MorphoGraphX analysiert und Zellteilungsmuster in einem wachsenden, zellulären 3D-Modell simuliert (Abb. 4). Uns interessierte, ob eine Standardregel die Teilungsmuster eines pflanzlichen Embryos erklären kann, der aus einer einzelnen kugelförmigen Zelle hevorgeht und an den Suspensor gebunden ist. Die Regel der kürzesten Wand bestätigt sich in der Tat für die ersten drei Teilungsrunden, da die Einzelzelle symmetrisch in acht Zellen unterteilt wird. Beim Übergang vom 8- zum 16-Zellen-Zustand weichen die Teilungen jedoch von dieser Regel ab. Jetzt erfolgen die bekannten querverlaufenden Teilungen, aus denen das Protoderm hervorgeht.


Ein weiteres Ergebnis unserer Untersuchungen war, dass mutante Embryonen mit unzureichender Auxin-Signalübertragung zu einer Teilung nach der Regel der kürzesten Wand zurückkehren. Auxine sind Pflanzenhormone, die Wachstum und Zellteilung anregen und steuern. Dies lässt vermuten, dass eine nicht standardmäßige Zellteilung speziell reguliert werden muss. Zudem fanden wir heraus, dass diese nicht standardmäßigen, regulierten Teilungen Tochterzellen erzeugen, die asymmetrisch sind. Solche asymmetrischen Teilungen sind oft ein Hinweis auf das Differenzierungsschicksal der Tochterzellen, in diesem Fall wäre es die Spezifizierung zur Bildung der äußeren Protodermzellen. Der ursächliche Zusammenhang zwischen geometrischer Asymmetrie und der asymmetrischen Identität der Tochterzellen ist derzeit unbekannt. Plausibel scheint jedoch die Annahme, dass sich die Zellgröße auf Genexpressionsmuster auswirken könnte.
Mechanische Modelle von Pflanzenzellen und Organen
Ein Großteil unserer Forschungsarbeit bezieht sich auf mechanische Modelle von Pflanzenzellen. Indentationsexperimente mit der CFM-Technik sind auf ein Simulationsmodell angewiesen, mit dem sich die Ergebnisse interpretieren lassen, da die gemessene Kraft eine Kombination aus Innenwandspannung, Zellwanddicke und Systemgeometrie repräsentiert. Benötigt wird ein quantitatives mechanisches Modell der Zelle, um die technischen Parameter zu reversieren, die wahrscheinlich einer genetischen Kontrolle unterliegen. Diese Daten werden in Simulationsmodelle des Pflanzenzellwachstums eingespeist, in denen Gene die physikalischen Eigenschaften der Zellen regulieren. Die in solchen Modellen entstehende Form resultiert aus der Interaktion zwischen Genetik und Mechanik [8]. Unsere frühen Arbeiten auf diesem Gebiet, die wir zusammen mit dem Bassel-Labor durchgeführt haben, lassen vermuten, dass das beobachtete Wachstum nicht nur ein direkter Readout der wachstumsfördernden Genexpression ist, sondern die zelluläre 3D-Struktur des Gewebes selbst von Bedeutung sein könnte. Größe und Anordnung der Zellen können sich enorm auf ihre Fähigkeit auswirken, sich auszudehnen und auf Wachstumsfaktorgradienten zu reagieren. Dies ist ein weiterer Mechanismus, über den mechanische Signale, die über die lokale Zellgeometrie des Pflanzengewebes ausgelöst werden, auf die Wachstumsregulierung einwirken können. Darüber hinaus wurde kürzlich nachgewiesen, dass kortikale Mikrotubuli - Bestandteile des Zytoskeletts - als Reaktion auf Belastungen ihre Ausrichtung verändern [9]. Und da die Mikrotubuli die Zellulose-Synthase-Komplexe steuern, kann deren Ausrichtung Einfluss auf die Anisotropie der Zellwand nehmen und auf die Zellform zurückwirken. Für die Aufklärung dieser Phänomene ist ein 3D-Zellmodell erforderlich, das sowohl die genetischen Regulierungsmechanismen als auch die Signalwege sowie deren Interaktionen und Rückwirkungen auf die physikalischen Prozesse von Wachstum und Mechanik umfasst. Und schließlich muss dieses Modell quantitativ sein, damit die Hypothese über die Mechanismen der Morphogenese experimentell überprüft werden kann.